
Roles of amacrine
cells
[General
characteristics] [Amacrine cell
circuitry as revealed by EM]
[A2] [AII] [A8] [A13] [A17] [A19 and A20] [A22] [A18] [Starburst cell] [References]

6. A13: a small-field diffuse amacrine cell of the cone system.
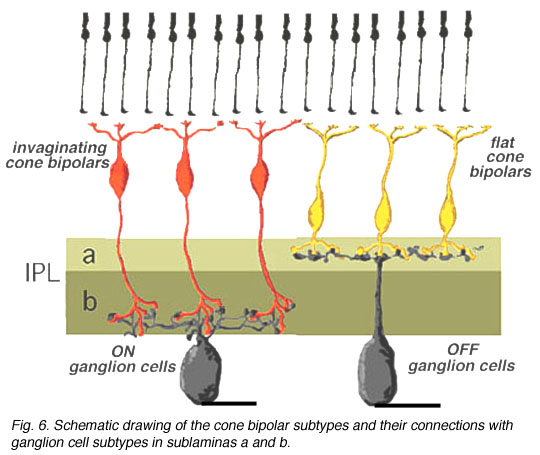
Cell A13 is a diffusely branched cell with a large cell body (12 um diameter) and fine dendrites bearing distinct beads at regular intervals that run through mostly strata 3-5 of sublamina b of the IPL to end up along the top of ganglion cell bodies. The complete tree covers a 100 um area.

Fig. 18. HRP and Golgi appearance of A13 amacrine cells (59 K jpeg image)
Electron microscopy shows that A13 cells, like A8 cells, get only a minor rod-bipolar-cell (12%) input while three different types of cone bipolar cell have major synaptic input (28%). The cone bipolar inputs are from axons in sublamina a, stratum 3-4 of sublamina b and stratum 5 of sublamina b. A13 cells appear to make reciprocal synapses upon cone bipolar cells and possibly also upon rod bipolar cells. Amacrine cells provide much more synaptic input than bipolar cells (60%). A13 makes synaptic output to OFF-center ganglion cells, of both alpha and beta types, in sublamina a. Gap junctions link A13 cells at their beaded dendrites (Kolb and Nelson, 1996).
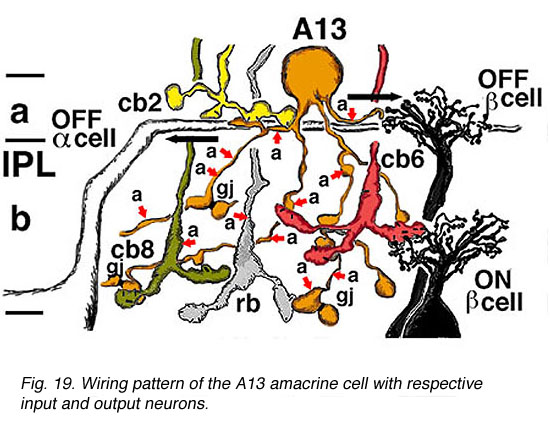
Fig. 19. Wiring pattern of the A13 amacrine cell (78 K jpeg image)
Click here to see an animation of the wiring pattern of the A13 amacrine
cell
(78 K quicktime movie)
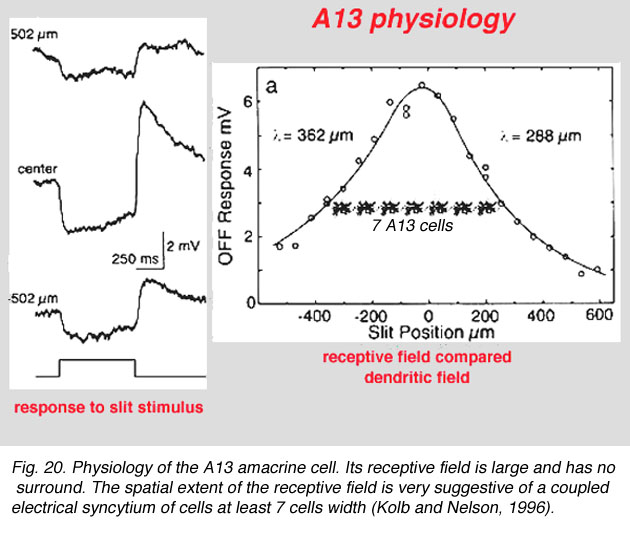
Fig. 20. Physiology of the A13 amacrine cell (59 K jpeg image)
7. A17: the wide-field reciprocal rod amacrine cell.
A17 is a wide-field diffuse amacrine. Its dendritic tree can span close to a millimeter of retinal surface. Its very fine dendrites bear pronounced beads at regular intervals along their length. The majority of the dendritic tree runs in sublamina b of the IPL, along the top of the ganglion cell layer in stratum 5 as a dense network of fine fibers. Over 1000 beads have been counted on such A17 amacrine cells in the cat retina (Nelson and Kolb, 1985), and, as we shall see later, the beads are the synaptic points where reciprocal synapses with rod bipolar cells occur (Sandell et al., 1987).
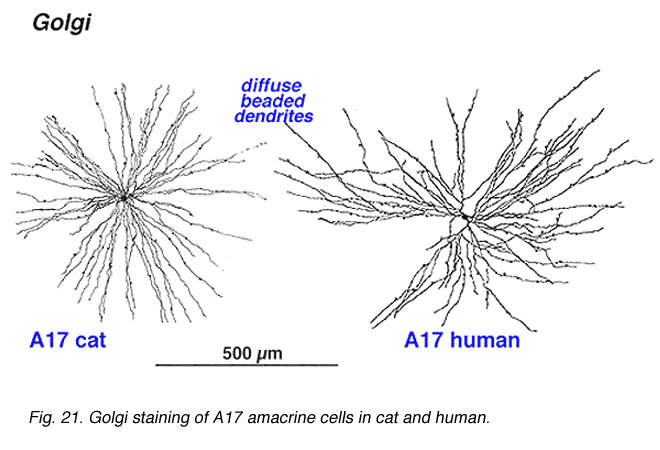
Fig. 21. Golgi staining of A17 amacrine cells (78 K jpeg image)
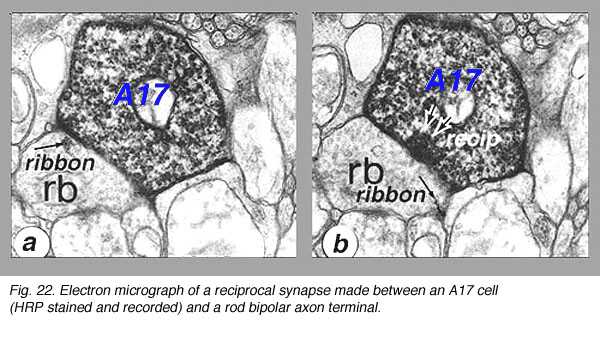
Electron microscopy shows that A17 cells are the predominant reciprocal amacrine cell at the rod bipolar cell axon terminals in sublamina b of the IPL. In addition their fine dendrites in sublamina a receive some synapses from the dopaminergic amacrine cell A18 (see below). However, there has not yet been discovered any synaptic output of the A17 except the reciprocal synapses with the rod bipolar axon.
Fig. 22. Electron micrograph of reciprocal synapse (78 K jpeg image)

8. A19 and A20: ON-OFF wide-field amacrine cells.
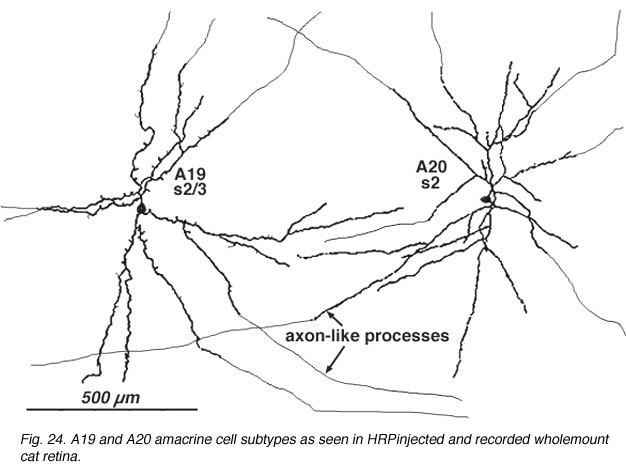
A19 and A20 are similar appearing wide-field radiate amacrine cells with 300-500 um diameter fields. However, their fields are further expanded to several mm by the presence of fine axon-like processes coming either from the ends of tapering dendrites (A19, A20) or from primary dendrites or even from the cell body.
Fig. 24. A19 and A20 amacrine cell subtypes (59 K jpeg image)

A19 and A20 are both transient depolarizing ON-OFF cells otherwise known as ON-OFF amacrine cells in response type. Such cells have been difficult to record long term responses from in a mammalian retina (Freed et al., 1996) and the example illustrated below of an A19 is one of the longest held and thus a receptive field was mapped.
Fig. 25. Physiological responses of the A20 amacrine cell (59 K jpeg image)
The receptive fields are large i.e >550 um radius, rather larger than their immediate dendritic tree sizes, and undoubtedly due to their axon-like extensions (Freed et al., 1996). Neither A19 nor A20 exhibited antagonistic surrounds.
Electron microscopy for circuitry of A19 and 22 has been performed. A19 receives two types of cone bipolar input (24%): the first is from OFF-center cb2 cone bipolar cells of sublamina a, while the other is ON-center input from cb5 at the sublamina a/b border where cb5 passes into sublamina b. OFF- and ON-center bipolar input could account for this cells ON-OFF physiology. The predominant input is from amacrine cells of sublamina a though (76%). Some of the amacrine inputs could be from rod amacrine AII cells, others from A2 cells while some looked like they could be from other A19 cells. A19's output is reciprocal synapses to both types of bipolar (40%), to unknown dark amacrines (40%) and possibly some fine diameter ganglion cell dendrites (20%) (Freed et al., 1996).
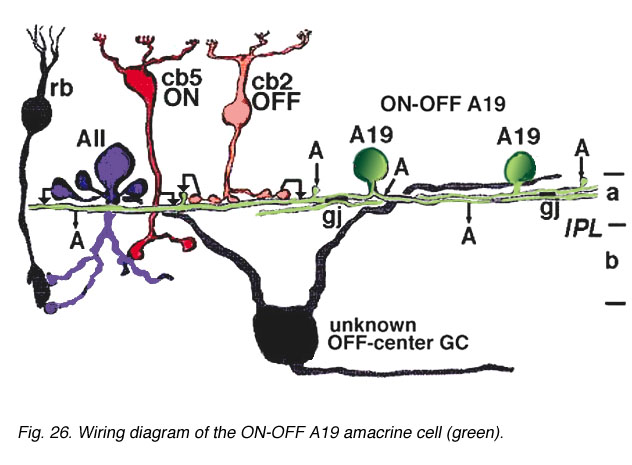
Fig. 26. Wiring diagram of the ON-OFF A19 amacrine cell (59 K jpeg image)
Gap junctions may connect A19 dendrites one to another (Fig. 11, A19, gj). A19 is GABA-ergic (Pourcho and Goebel, 1983).
A20 cells, running in upper stratum 2 of sublamina a have slightly different synaptic relationships with other nerve cells in the neuropil, from A19, which will be remembered, sits on the sublamina a/b border (above). Thus A20 receives some cone bipolar inputs in stratum 1/2 and 2 (7%) but they are dominated by amacrine cell inputs (93%). Much of this input is recognizable as being from lobular appendages of rod AII cells. A20's output is about equally as reciprocal synapses to bipolar cells (5%) and synapses to amacrine cells (5%), but the cells have major output to Alpha ganglion cells of sublamina a (90%). The latter ganglion cells would be OFF center types of course.

Fig. 27. Wiring diagram of the ON-OFF A20 amacrine cell (59 K jpeg image)
Both A19 and A20 cells are probably involved in the transfer of fast messages from one area of retina to another. They may be the basis of the proximal negative response recordable in the intraretinal ERG and in the shift effect in Y type ganglion cells.
9. A22: A putative substance P containing, ON-OFF neuron of the cone system.
Immunocytochemical staining with antibodies against the neuropeptide substance P (SP) reveal a large-field amacrine cell in human retina (Cuenca et al., 1994). The SP containing amacrine is possibly equivalent to the wide-field A22 cell of cat that has been intracellularly recorded as an ON-OFF cell (see above) (Freed et al., 1996).
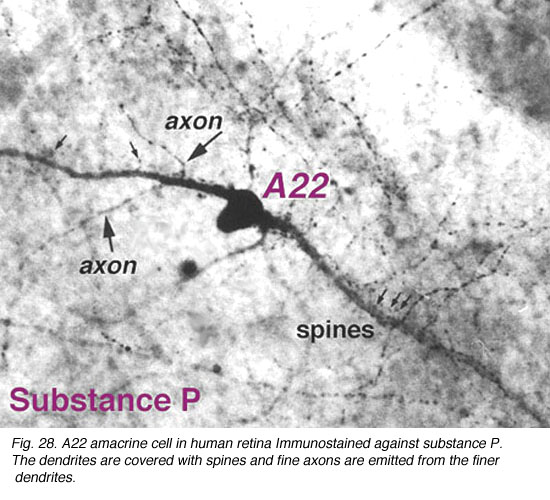
SP-containing and A22 cells are wide-field amacrine cells (dendritic
trees of 500 um span), often having large cell bodies (14-16 um) displaced to
the ganglion cell layer, with their major dendritic stratification in strata 3
and 4 of sublamina b of the IPL. Both cells have long axon-like processes
that pass up into sublamina a to run for up to a mm and also down into
stratum to run over ganglion cell bodies. Their major dendrites are covered with
spines.
Fig. 28. Substance P containing amacrine cell in human retina (59 K jpeg image)
Electron microscopy shows that the spines are postsynaptic with reciprocal synapses to cone bipolar cells. They also receive synapses from unidentified amacrine cells and other SP-IR cell dendrites. They are presynaptic to ganglion cell dendrites and directly to ganglion cell bodies (possibly some of the latter are of SP-IR ganglion cells). Their axonal processes are presynaptic to amacrine and ganglion cells in stratum 5 and in stratum 1 of the IPL. Probably the latter ganglion cells are OFF-center (see above). They receive cone bipolar input from a cone bipolar type in stratum 1 of the IPL. Almost certainly A22 cells, SP-IR amacrine cells of cat and human are GABA-ergic (Porcho and Goebel, 1988). This ON-OFF cell probably plays a similar role as the A19 and A20 cells above in being recordable in the proximal negative response (PNR) (Burkhardt, 1970) and being involved in fast moving aspects of visual coding.
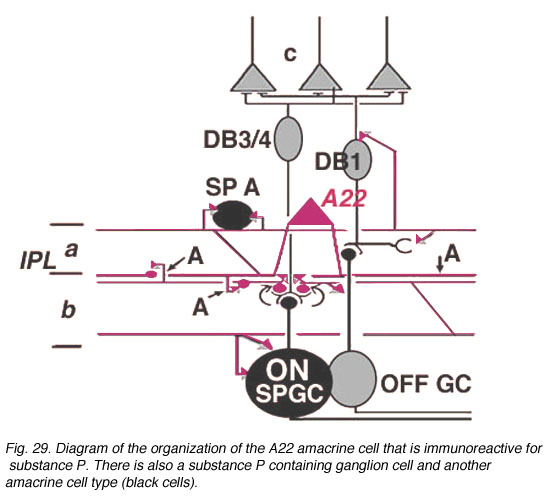
Fig. 29. Diagram of the organization of the A22 amacrine cell (59 K jpeg image)
[General
characteristics] [Amacrine cell
circuitry as revealed by EM]
[A2] [AII] [A8] [A13] [A17] [A19 and A20] [A22] [A18] [Starburst cell] [References]
.gif)
![]()
![]()
![]()
{kind=link}